Material sobre plantas




1) História da classificação das plantas:
As plantas, como tendo papéis fundamentais na vida do homem, seja na alimentação ou no uso medicinal, sempre chamaram a atenção do homem como um ser investigativo e curioso. Assim, as primeiras tentativas de registrar as plantas e classificá-las datam desde a Antiguidade, com documentos escritos por diversas civilizações (China, Egito, povos mesopotâmios, etc.), sendo que na Grécia Antiga seriam encontradas as principais obras, até então, abordando o estudo das plantas, grande parte delas de autoria do filósofo Teofrasto, considerado como "o pai da botânica". Os trabalhos do sábio grego seriam conservados pelas obras de Plínio ("Historia Naturalis") e de Discórides, que se dedicou a estudar principalmente o uso medicinal das plantas em sua obra "Materia Medica", repleto de descrições e esquemas destas plantas.
 |
| Teofrasto (372 a.C. - 287 a.C.) |


Na Idade Média, se deu prosseguimento ao estudo das ervas medicinais (herbalismo), muito baseado nas obras de Dioscórides, produzidas séculos atrás. Devido à precária tecnologia disponível na época, análises muito profundas da anatomia das plantas era incabível. Mesmo assim, de vez em quando eram publicadas algumas novas descobertas desta área, como a diferenciação da estrutura do caule das plantas monocotiledôneas e dicotiledôneas pelo filósofo alemão Albertus Magnus. Enquanto as ervas eram comumente classificadas de acordo com as suas propriedades específicas, como na obra "Herbarum vivae eicones" de Otto Brunfels, outros botânicos, como Jerome Block, que classificou centenas de plantas em arbustos, ervas e árvores. Classificação simples, mas que era mais adequada aos limites do conhecimento e da tecnologia da época.
 |
| Albertus Magnus (1200-1280) |

Além de levar em consideração as diferenças de tamanho e aparência, o cientista Andrea Cesalpino organizou, em seu "Livro das plantas" ("De plantis libri"), centenas de espécies com base nas flores, frutos e sementes que apresentavam. É importante ressaltar que a sua classificação ainda incluía os fungos e as algas marinhas, consideradas como plantas pela comunidade científica da época.
 |
| Andrea Cesalpino (1519-1603) |
Em seus trabalhos, os irmãos Jean e Gaspard Bauhin também trouxeram importantes avanços para a classificação das plantas com a introdução de um sistema de nomenclatura binomial, o qual seria utilizado por Lineu, nome de peso no ramo da sistemática. Com as novas descobertas científicas decorrente do aprimoramento do microscópio por van Leeuwenhoek, como a descoberta das células, a botânica se distanciou de outras áreas, como o herbalismo, embora mantivesse as suas contribuições. Os critérios utilizados para classificar as plantas se tornaram cada vez mais elaborados. Em seus trabalhos, o inglês John Ray classificou as plantas em imperfeitas e perfeitas. As plantas imperfeitas reuniam as criptógamas, isto é, que não apresentam sementes (e, como consequência, também não apresentam flores e frutos). Já as perfeitas correspondem aos espermatófitos, que se reproduzem por meio de sementes, algumas desenvolvendo frutos e flores. As plantas perfeitas também eram divididas em monocotiledôneas e dicotiledôneas.

Mesmo tendo sido introduzido muito tempo antes, o sistema binomial de nomenclatura ganharia maior reconhecimento com Lineu e sua obra "Systema Naturae". Seus novos conceitos de espécie e de hierarquia taxonômica ajudariam a organizar melhor a diversidade das plantas. Lineu também foi um dos primeiros a identificar a espécie de uma planta com base nas estruturas de suas flores.
 |
| Carl von Linné (Lineu) (1707-1778) |

Depois de Lineu, a introdução dos conceitos de evolução por Charles Darwin seria o próximo impacto na taxonomia das plantas, pois se buscou organizar as plantas das formas mais "primitivas" até as mais "complexas", formando-se, então, relações de parentesco genético entre esses grupos (filogenia), o que se firma com as descobertas de Gregor Mendel no campo da genética.
A primeira classificação taxonômica das plantas que seguiu tal preceito foi a de August W. Eichler. O seu sistema, publicado em 1883, dividia as plantas em não produtoras de sementes (criptógamas) e produtoras de sementes (fanerógamas). Dentro das criptógamas incluía algas, fungos, briófitas e pteridófitas e, nas fanerógamas, incluía gimnospermas e angiospermas, as últimas, por sua vez, eram divididas em mono e dicotiledôneas. Pode-se dizer que o sistema de Eichler, apesar da inclusão de algumas espécies já não mais consideradas como plantas atualmente (algas e fungos), é praticamente similar à classificação utilizada atualmente.

Com base no sistema de Eichler, seria desenvolvido pelos botânicos Adolf Engler e Karl Prantl, um outro sistema muito similar, mas este sim abordando os diferentes grupos com base em seus graus evolutivos.


2) Características gerais das plantas: as plantas são as espécies que pertencem ao reino Plantae (ou, também, Metaphyta) do domínio Eukarya. Se distinguem dos demais seres eucariontes por causa dos seguintes traços:

a) São seres exclusivamente multicelulares, dotados de parede celular (principal substância componente = celulose) e de cloroplastos, organelas que armazenam a clorofila.


b) Realizam a fotossíntese por meio de seus cloroplastos, produzindo açúcares (glicose) e oxigênio a partir de gás carbônico, água e luz. Possuem nutrição autotrófica, então, pois produzem seu próprio alimento. Sua reserva energética é o amido.

c) Apresentam uma alternância de gerações assexuada e sexuada ao longo de seus ciclos de vida (metagênese). Fase assexuada: esporofítica - produz esporos para se reproduzir, o esporo dá origem ao gametófito. Fase sexuada: gametofítica - produz gametas que, ao se encontrarem, formam um zigoto, que se desenvolve na forma de um esporófito. Plantas de diferentes grupos possuem fases predominantes distintas.

a) São seres exclusivamente multicelulares, dotados de parede celular (principal substância componente = celulose) e de cloroplastos, organelas que armazenam a clorofila.

b) Realizam a fotossíntese por meio de seus cloroplastos, produzindo açúcares (glicose) e oxigênio a partir de gás carbônico, água e luz. Possuem nutrição autotrófica, então, pois produzem seu próprio alimento. Sua reserva energética é o amido.

c) Apresentam uma alternância de gerações assexuada e sexuada ao longo de seus ciclos de vida (metagênese). Fase assexuada: esporofítica - produz esporos para se reproduzir, o esporo dá origem ao gametófito. Fase sexuada: gametofítica - produz gametas que, ao se encontrarem, formam um zigoto, que se desenvolve na forma de um esporófito. Plantas de diferentes grupos possuem fases predominantes distintas.

d) Vivem principalmente na terra firme. Algumas espécies, como os musgos, por exemplo, vivem em ambientes mais próximos da água ou onde há um grau de umidade considerável.
e) Possuem tecidos e órgãos específicos para determinadas tarefas, como os tecidos que formam os canais de condução de seiva, por exemplo.
3) Estrutura celular:


Sendo seres eucariontes, as plantas apresentam um elaborado sistema de organelas no interior de suas células, além de estruturas que são características destes organismos.
a) O cloroplasto: é a organela responsável por realizar a fotossíntese das plantas. Se trata de um dos vários tipos de plastos existentes nas plantas, se destacando por armazenar a clorofila, substância que confere a coloração verde às plantas. Os cloroplastos apresentam uma estrutura complexa, formada por duas membranas, uma externa e uma interna (entre elas está o espaço intermembranoso). A membrana interna delimita o espaço interno dos cloroplastos, onde se encontram várias grana de tilacoides (um granum = uma pilha de tilacoides, grana = conjunto de granum). Os tilacoides são espécies de "sacos" em formato de discos que guardam a clorofila. Os granum de tilacoides são organizados em espécies de "prateleiras", as lamelas e o restante do espaço interno é preenchido por uma matriz chamada estroma. Assim como as mitocôndrias, os cloroplastos também possuem seu próprio DNA, o que fortalece a teoria de que ambas as organelas eram microrganismos que acabaram sendo fagocitados por uma célula eucarionte maior, formando uma relação simbiótica em que fornecem energia para a célula enquanto esta garante a proteção do microrganismo (teoria de endossimbiose).


c) Vacúolo: são vesículas preenchidas de fluido que podem variar de tamanho à necessidade da célula. Quando a célula absorve água em excesso, por exemplo, o vacúolo recebe grande parte desta água e expande. O vacúolo, então, também tem função de armazenar substâncias de forma geral.
4) Nutrição e metabolismo: as plantas, como já foi falado, obtêm sua alimentação por meio da fotossíntese a partir de água, gás carbônico e luz. Os açúcares resultantes deste processo podem ser armazenados como reserva na forma de amido. Além da fotossíntese, as células vegetais são dotadas de mitocôndrias, assim, usam o açúcar produzido na sua fotossíntese para obter energia, que fica guardada nas moléculas de ATP através da respiração celular. Como a fotossíntese é o processo mais característico das plantas, faremos uma descrição mais aprofundada deste processo, e não da respiração celular.
A fotossíntese:
Absorção de água: as plantas conseguem a água, que cumpre diversas funções nelas além da fotossíntese, como diminuir a temperatura da planta através da evaporação; conduzir nutrientes e controlar a turgidez das células, etc. por meio de suas raízes. Absorvendo a água encontrada no solo, esta é conduzida por meio dos sistemas vasculares (nas plantas vasculares) ou por difusão, de célula em célula, nas briófitas.


Absorção de luz: o pigmento armazenado nos cloroplastos, a clorofila (do tipo a ou b) absorvem partes específicas da radiação solar, mais especificamente, as partes correspondente aos espectros vermelho e azul do espectro visível, assim, refletem as ondas do espectro verde o que faz com que as plantas adquiram a sua coloração característica. Outros pigmentos que podem participar da fotossíntese em algumas espécies, como a cenoura, são os carotenoides, que não absorvem as ondas mais avermelhadas, logo, têm uma coloração com esses tons (amarelo, laranja ou vermelho). A quebra dos pigmentos durante a estação do outono é responsável pela mudança de cor das folhas (de verde para um tom mais marrom-avermelhado).





A fotofosforilação acíclica envolve ambos os fotossistemas I e II. Ao captar a energia solar, o fotossistema II perde um par de elétrons, que são transferidos para o fotossistema I por meio de uma cadeia transportadora de elétrons. Ao longo deste percurso, a energia perdida é utilizada para realizar a fosforilação do ADP, formando o tão desejado ATP. Um dos processos característicos deste tipo de fosforilação é a quebra das moléculas de água em gás oxigênio e prótons (H+). O gás oxigênio deixa a célula e os prótons são utilizados na redução do NADP+ em NADPH, que funciona, praticamente, como um transportador de prótons no processo de fotossíntese. A energia utilizada para esta redução provém dos elétrons perdidos pelo fotossistema I, estes que são repostos pelo par de elétrons originalmente pertencentes ao fotossistema II.
A fotossíntese:
Absorção de água: as plantas conseguem a água, que cumpre diversas funções nelas além da fotossíntese, como diminuir a temperatura da planta através da evaporação; conduzir nutrientes e controlar a turgidez das células, etc. por meio de suas raízes. Absorvendo a água encontrada no solo, esta é conduzida por meio dos sistemas vasculares (nas plantas vasculares) ou por difusão, de célula em célula, nas briófitas.
Entrada e saída de gases: as folhas das plantas possuem estruturas dedicadas à entrada e saída do dióxido de carbono (CO2) e oxigênio (O2), respectivamente, os estômatos. Os estômatos abrem durante o dia através de um processo complexo em que a concentração de íons no interior de suas células-guarda aumenta. Assim, recebem grandes quantidades de água através da osmose, aumentando a sua turgidez. Com o aumento da turgidez, as células assumem um formato "curvado", abrindo um poro pelo qual os gases entram ou saem. O fechamento dos estômatos geralmente ocorre durante a noite, em que as células-guarda se desfazem dos íons absorvidos durante a manhã, levando à perda da turgidez, retornando ao seu formato original. Além de fecharem durante o período noturno, os estômatos também podem se cerrar para evitar a perda de água por meio da transpiração.




Fase luminosa/clara: a primeira fase da fotossíntese ocorre no interior dos tilacóides, ou melhor, em suas membranas. As membranas dos tilacóides apresentam várias proteínas dedicadas a este processo, entre elas, os principais são os fotossistemas I e II. O objetivo final dessa fase é a síntese de ATP, que vai ser utilizada na fase seguinte para a produção da glicose. Em geral, existem duas vias pelas quais este objetivo pode ser atingido: a fotofosforilação (produção de ATP a partir de energia luminosa) cíclica ou acíclica.
Na fotofosforilação cíclica, só é envolvido o fotossistema I. Quando este complexo absorve a energia solar na forma de fótons, ele entra em um estado de "agitação" e perde um par de elétrons. Estes elétrons são transferidos por meio de uma série de proteínas (cadeia de transporte de elétrons) até retornarem ao fotossistema I, de onde saíram. Como os elétrons "perdidos" retornam ao seu local de origem, este processo é conhecido como "cíclico". Porém, qual o objetivo de todo este processo se, no final dele, tudo volta a como estava antes? Durante este trajeto de proteína em proteína, os elétrons vão ficando menos energizados, assim, esta energia liberada pode ser usada para outros fins, o que, neste caso, é a produção do ATP. Para entender melhor esta perda de energia, lembro que, nas aulas de biologia do 1o ano do ano passado, o professor Fábio comparou este processo à brincadeira da "batata quente", porque enquanto a batata passa de mão em mão na roda, ela vai esfriando. Esta mesma lógica pode ser aplicada no caso da cadeia de transporte de elétrons, pois assim que passam de proteína em proteína, os elétrons vão perdendo energia.

A fotofosforilação acíclica envolve ambos os fotossistemas I e II. Ao captar a energia solar, o fotossistema II perde um par de elétrons, que são transferidos para o fotossistema I por meio de uma cadeia transportadora de elétrons. Ao longo deste percurso, a energia perdida é utilizada para realizar a fosforilação do ADP, formando o tão desejado ATP. Um dos processos característicos deste tipo de fosforilação é a quebra das moléculas de água em gás oxigênio e prótons (H+). O gás oxigênio deixa a célula e os prótons são utilizados na redução do NADP+ em NADPH, que funciona, praticamente, como um transportador de prótons no processo de fotossíntese. A energia utilizada para esta redução provém dos elétrons perdidos pelo fotossistema I, estes que são repostos pelo par de elétrons originalmente pertencentes ao fotossistema II.

Em suma: na fase luminosa, começamos com água e energia luminosa e terminamos com gás oxigênio (que deixa a célula por difusão), moléculas transportadoras de hidrogênio (na forma de prótons) e ATP.
Fase escura: a segunda e última fase da fotossíntese, ocorre independentemente da "oferta" de luz ou não. Partindo de onde paramos, ainda temos, no cloroplasto, os transportadores de hidrogênio e algumas moléculas de ATP. Nesta fase, também entra o gás carbônico e o processo como um todo é realizado na matriz que preenche o cloroplasto, isto é, o estroma. Ou seja, ao contrário da fase luminosa, esta acontece fora dos tilacóides. A fase escura, em si, consiste na reorganização de todos estes compostos e substâncias por meio de um processo chamado Ciclo de Calvin:

Basicamente, no Ciclo de Calvin, os hidrogênios são transferidos dos transportadores de prótons para reagir com as moléculas de gás carbônico, com a energia do ATP servindo para reorganizar as ligações e formar as moléculas de glicose, que podem ser utilizadas pelas mitocôndrias para sintetizar mais moléculas de ATP.



A respiração celular utiliza os produtos sintetizados pela fotossíntese, mais especificamente o gás oxigênio e a glicose, para, através de uma série de reações químicas, gerar energia que a planta pode utilizar para realizar as suas funções e crescer. A principal organela envolvida neste processo é a mitocôndria que, assim como os cloroplastos, acredita-se que tenha se originado a partir de uma relação simbiótica entre uma célula eucarionte e uma bactéria fagocitada há muito tempo atrás.

A respiração tem início com a glicólise, a quebra da molécula de açúcar, a glicose, ainda no citoplasma celular. Esta fase não precisa, necessariamente, da presença de oxigênio para ocorrer, logo, ela pode acontecer ainda em ambientes anaeróbicos. Na glicólise, duas moléculas de ATP são dissolvidas, incrementando grupos fosfato à molécula (fosforilação) e tornando-a mais instável, facilitando a sua quebra. A glicose, então, é oxidada, liberando energia suficiente para a formação de quatro moléculas de ATP (total ganho = 2 moléculas de ATP), duas moléculas de NADH (moléculas transportadoras de prótons/hidrogênio), além de duas moléculas orgânicas de três carbonos cada, o piruvato (ou ácido pirúvico).


Resumo da glicólise:
Consumo: 1 molécula de glicose e 2 moléculas de ATP
Produção: 2 moléculas de piruvato, 2 moléculas de NADH e 4 moléculas de ATP
Rendimento: 2 moléculas de piruvato, 2 moléculas de NADH e 2 moléculas de ATP
Antes de iniciar o ciclo de Krebs, as moléculas de piruvato precisam passar por mais uma série de reações (estas já no interior da mitocôndria) a fim de serem convertidas em Acetil-CoA (Acetil coenzima A), sendo esta fase conhecida como a da "oxidação do piruvato". Inicialmente, um grupo carboxila (-COOH) é removido do piruvato e liberado na forma de gás carbônico (este processo ocorre em ambas aas moléculas de piruvato resultantes da glicólise, ou seja, são liberados duas moléculas de gás carbônico ao todo). Desta maneira, cada molécula de piruvato, com três carbonos, passa para uma molécula de dois carbonos. Estas moléculas resultantes são oxidadas, formando o acetato enquanto os seus elétrons são recolhidos, formando uma molécula de NADH por cada acetato (ou seja, se formam dois NADH) e, por fim, este acetato é associado à coenzima A, composto derivado da vitamina B, formando Acetil coenzima A.
Resumo da oxidação do piruvato:
Consumo: 2 moléculas de piruvato (C3H4O3)
Produção: 2 acetila (C2H3O) associados à coenzima A (acetil-CoA), 2 moléculas de gás carbônico (CO2) e 2 NADH


Em seguida, as moléculas de acetil-CoA passam, ainda no interior da matriz mitocondrial, por um processo de oito etapas denominado "ciclo de Krebs":
1- Os acetil-CoA são combinados com moléculas de quatro carbonos (oxaloacetato), formando substâncias com seis carbonos (citratos)
2 - Um átomo de hidrogênio e um agrupamento hidroxila (-OH) são removidos dos citratos e reposicionados na molécula, formando o isocitrato (ainda com seis carbonos)
3 e 4 - Nas etapas 3 e 4 ocorre a perda de um carbono na forma de gás carbônico além da formação de um NADH, o resultado é uma molécula de quatro carbonos chamada alfa-cetoglutarato. Na etapa 4 ocorre o mesmo (desprendimento de dióxido de carbono, levando o número de carbonos para quatro, assim como mais um NADH). O resultado da etapa 4 é uma molécula de quatro carbonos que se liga a uma coenzima A, formando o complexo succinil-coenzima A.
5 - A coenzima A do composto é substituída por um grupo fosfato, que é liberado para a formação de uma molécula de ATP. O resultado é o succinato, que mantém os seus quatro carbonos.
6 - O succinato é oxidado e se desfaz de dois átomos de hidrogênio que são recolhidos pelo complexo FAD, resultando no FADH2. Em decorrência desta oxidação, o succinato é convertido em fumarato, também com quatro carbonos.
7 - É incrementada uma molécula de água ao fumarato, formando o malato, ainda uma molécula de quatro carbonos.
8 - Finalmente, o malato se desfaz de um elétron, que é carregado pelo NADH. Desta maneira, o malato é convertido em oxalacetato novamente, que será utilizado no próximo ciclo de Krebs.

Ao todo, com dois complexos de acetil-coenzima A, são produzidos 6 NADH (3 por ciclo), 2 FADH2 (1 por ciclo), 2 ATPs (1 por ciclo) e 4 CO2 (2 por ciclo)
A última etapa da respiração celular é a fosforilação oxidativa, em que o grosso das moléculas de ATP são produzidas. A fosforilação oxidativa ocorre na interna da mitocôndria, que separa as suas matrizes interna e externa. Esta membrana interna possui uma série de proteínas que compõem uma cadeia de transporte de elétrons (algo parecido com o que ocorre nos cloroplastos, a proteína recebe os elétrons e os passam para as proteínas seguintes, sendo que, durante este processo, os elétrons vão perdendo energia). Neste caso, quem fornece os elétrons para a cadeia de transporte são as moléculas carregadoras de elétrons (os NADH e FADH2) produzidas até então. Estas moléculas, portanto, cedem os elétrons a estas proteínas, que passam os elétrons adiante até chegar no aceptor final, o oxigênio. Ao receber os elétrons, o oxigênio é reduzido e forma água. Os íons de hidrogênio que transferiam estes elétrons para os citocromos então atravessam a membrana e criam uma diferença de potencial entre os meios interno e externo, estabelecendo uma forma de acúmulo de energia que é liberado para formar o ATP a partir do ADP (adenosina difosfato) e Pi (fosfato inorgânico). De forma geral, foram sintetizados 10 NADH (2 da glicólise, 2 da oxidação do piruvato e 6 do ciclo de Krebs) e 2 FADH2 ao longo do processo de respiração celular, assim, podem ser formados, somente na fosforilação oxidativa, 34 ATPS (30 dos NADH, mais energéticos e 4 do FADH2, menos energéticos).


Em suma, na fosforilação oxidativa, é formado água e 34 moléculas de ATP a partir dos carregadores de elétrons.
O quadro geral da respiração celular é, então, que a partir de uma molécula de glicose, são produzidos:
- 6 moléculas de CO2 (2 da oxidação do piruvato e 4 do ciclo de Krebs)
- 6 moléculas de H2O (oriundos da fosforilação oxidativa)
- saldo de 38 moléculas de ATP (2 da glicólise, 2 do ciclo de Krebs e 34 da fosforilação oxidativa)


Como as plantas realizam tanto a fotossíntese quanto a respiração aeróbica, parte do gás oxigênio e da glicose que produzem é consumida pela planta para a produção de energia.
Diferentemente da fotossíntese, a respiração celular não depende da intensidade de luz absorvida, podendo acontecer em qualquer horário, tanto de noite quanto de dia.
A intensidade da fotossíntese depende de quanta luz a planta absorve. Quanto mais escuro o ambiente, menos "produtiva" será a fotossíntese (menos açúcares e gás oxigênio serão sintetizados) e vice-versa. Quando a planta produz mais matéria orgânica que precisa para suprir as suas necessidades energéticas, ela consegue utilizar essa matéria orgânica para "investir" no seu crescimento.
Define-se, então, a intensidade luminosa que a planta precisa receber para compensar os seus "gastos" com a respiração celular através da fotossíntese, o Ponto de Compensação Luminosa (Ou Ponto de Compensação Fótico).
Absorvendo uma intensidade de luz igual ao seu "PCL", a planta não apresenta nem ganhos e nem perdas, pois todos os açúcares e gases produzidos por meio da fotossíntese acabam sendo utilizados para a geração de energia na respiração (o gás carbônico e a água liberados, por sua vez, também são totalmente consumidos na fotossíntese).
Ao absorver uma intensidade inferior ao ponto fótico, a planta produz menos do que a respiração celular demanda e, assim, ela não obtém os nutrientes necessários para poder gerar energia e muito menos crescer. Como a fotossíntese é reduzida nessa situação, a respiração celular se torna predominante e, dessa forma, mais gás carbônico é liberado do que gás oxigênio.
Já quando a planta recebe uma intensidade luminosa superior ao que é considerado o seu ponto de compensação, a fotossíntese acaba sendo intensificada e, consequentemente, são formados mais açúcares e gás oxigênio. Parte destes produtos são utilizados na respiração celular e o resto é usado para o crescimento.
A intensidade dos processos de geração de energia e nutrientes da planta em função da intensidade luminosa pode ser demonstrada na forma de gráfico, em que o encontro das linhas que representam a taxa de respiração celular e de fotossíntese (ou seja, o ponto em que há um consumo total dos gases resultantes destes processos) determina o seu ponto de compensação fótico:

O ponto fótico pode variar de planta para planta, assim, algumas espécies podem ser classificadas em conjuntos dependendo de quanta luz precisam absorver para compensarem as suas perdas gasosas. Aquelas plantas que requerem uma menor intensidade de luz, ou seja, têm um ponto de compensação fótico mais baixo, podem viver em áreas menos iluminadas, e, por isso, são chamadas de "umbrófilas" (umbra = sombra, portanto "plantas com afinidade por sombras"). Já as plantas que têm uma maior demanda de luz para compensar a respiração celular precisam viver em ambientes mais iluminados, sendo denominadas, então, "heliófilas" (helios = Sol, portanto "plantas com afinidade pelo Sol")

5) Classificação das plantas: quando falamos de plantas, geralmente estamos nos referindo às plantas terrestres, ou embriófitas, a fim de distingui-las de outros organismos, como as algas, que vivem em ambientes aquáticos e, algumas vezes, também são classificados como plantas. A apomorfia que origina o grupo das embriófitas é que seus organismos apresentam, em alguma parte de sua vida, embriões multicelulares maciços que se desenvolvem à custa do organismo materno.
As plantas comumente são divididas em 4 grupos principais: briófitas, pteridófitas, gimnospermas e angiospermas. Na verdade, estes termos são mais "generalizantes", pois cada um destes grupos inclui vários filos diferentes. Além disso, é comum agrupar alguns destes filos com base nas estruturas que apresentam. Primeiramente, se divide as plantas com base na presença ou ausência de vasos condutores de seiva, respectivamente as traqueófitas (que reúne pteridófitas, gimnospermas e angiospermas) e as briófitas. Depois, dentro das traqueófitas se encontram as espermatófitas, que apresentam semente, incluindo as gimnospermas e as angiospermas e, dentre as espermatófitas, se destacam as angiospermas, que apresentam frutos e flores.



a) Briófitas:
- São os seres mais primitivos dentre as embriófitas
- Não apresentam um sistema vascular para conduzir seiva
- Têm um porte consideravelmente pequeno quando comparado às demais plantas
- Apresenta, como fase dominante em seu ciclo de vida, o gametófito
- Se fixam ao subterrâneo por meio de raízes primitivas (rizoide) e algumas espécies desenvolvem caules ou folhas primitivas (cauloides e filoide respectivamente), portanto, não apresentam as formas verdadeiras destas estruturas
- Dependem de água para se reproduzir (o seu gameta masculino, o anterozoide é natante, ou seja, precisa nadar para fecundar o gameta feminino)
- Vivem em ambientes úmidos e sombreados
Classificação das briófitas: o termo "briófita" é utilizado generalizadamente para se referir a três filos distintos: Bryophyta (briófitas como os musgos), Anthocerophyta (antóceros) e Hepatophyta (hepáticas).
As plantas comumente são divididas em 4 grupos principais: briófitas, pteridófitas, gimnospermas e angiospermas. Na verdade, estes termos são mais "generalizantes", pois cada um destes grupos inclui vários filos diferentes. Além disso, é comum agrupar alguns destes filos com base nas estruturas que apresentam. Primeiramente, se divide as plantas com base na presença ou ausência de vasos condutores de seiva, respectivamente as traqueófitas (que reúne pteridófitas, gimnospermas e angiospermas) e as briófitas. Depois, dentro das traqueófitas se encontram as espermatófitas, que apresentam semente, incluindo as gimnospermas e as angiospermas e, dentre as espermatófitas, se destacam as angiospermas, que apresentam frutos e flores.


a) Briófitas:
- São os seres mais primitivos dentre as embriófitas
- Não apresentam um sistema vascular para conduzir seiva
- Têm um porte consideravelmente pequeno quando comparado às demais plantas
- Apresenta, como fase dominante em seu ciclo de vida, o gametófito
- Se fixam ao subterrâneo por meio de raízes primitivas (rizoide) e algumas espécies desenvolvem caules ou folhas primitivas (cauloides e filoide respectivamente), portanto, não apresentam as formas verdadeiras destas estruturas
- Dependem de água para se reproduzir (o seu gameta masculino, o anterozoide é natante, ou seja, precisa nadar para fecundar o gameta feminino)
- Vivem em ambientes úmidos e sombreados
Classificação das briófitas: o termo "briófita" é utilizado generalizadamente para se referir a três filos distintos: Bryophyta (briófitas como os musgos), Anthocerophyta (antóceros) e Hepatophyta (hepáticas).
 |
| Musgos |
 |
| Hepáticas |
 |
| Antóceros |
a) Musgos:
Seu gametófito haploide possui uma estrutura cauloide que se desenvolve perpendicularmente em relação ao solo, ao qual se fixa por meio de uma estrutura multicelular denominada rizoide, que desempenha a função de ancorar a planta ao substrato.

O gametófito do musgo tem simetria axial, isto é, se ele for partido ao meio, as duas metades serão simétricas.

Ao longo deste cauloide, também se formam estruturas que lembram folhas, os filoides. Estes filoides são os principais realizadores da fotossíntese nos musgos e, também, absorvem grande parte da água utilizada por estes organismos assim como os nutrientes e sais minerais que se encontram dissolvidos na água. Nos musgos, os filoides se dispõem em espiral ao redor do cauloide.
Os musgos masculinos e femininos possuem suas próprias estruturas reprodutivas, ambas se formando no topo do gametófito. A estrutura reprodutiva masculina é o anterídio, que constitui de vários sacos feitos de células mortas em que, em seus interiores, se encontram as células férteis, que formam os anterozoides, os gametas masculinos, que são dotados de motilidade, possuindo um par de flagelos. Os anterozoides são lançados ao exterior quando o anterídio rompe, permitindo que estes nadem até chegar ao arquegônio feminino. O arquegônio, ao contrário do anterídio, produz apenas um gameta por vez, que, no caso do musgo feminino, é o ovo (ou, também, oosfera). Além do ovo, o arquegônio apresenta uma estrutura em formato de tubo, o qual se preenche de fluido quando o arquegônio se encontra em sua fase madura, permitindo que os anterozoides nadem até a oosfera, fecundando-a. A fecundação dá origem a um zigoto diploide, o qual passa por uma série de mitoses, formando uma massa multicelular maciça chamada de embrião, que se mantém conectada à planta-mãe e se alimenta por meio dela através de uma estrutura chamada placenta. Após mais algumas divisões celulares, o embrião já começa a diferenciar suas estruturas, se implantando no arquegônio da mãe por meio do pé, e começa a crescer. Além do pé, se formam, também, outras duas partes principais: a seta e a cápsula. A seta é uma estrutura filamentosa por meio da qual se transferem os nutrientes do gametófito para o esporófito. No ápice da seta se encontra a cápsula, ou, também, esporângio, que contém os esporócitos diploides, que produzem os esporos haploides através da meiose. A cápsula em si também apresenta outras partes, dentre elas uma espécie de "tampa" ou "boca", o opérculo, além de uma cobertura que se desfaz quando o esporófito chega à maturidade e está pronto para liberar seus esporos, a caliptra. Os esporos, liberados, se implantam no substrato e germinam. Nos musgos, os esporos formam, primeiro, uma rede de filamentos chamada de protonema, o qual cresce, desenvolvendo as suas estruturas e dando origem ao gametófito e suas estruturas reprodutivas. Ao atingir a sua maturidade, os gametófitos já estão com suas estruturas reprodutivas bem desenvolvidas e capazes de reiniciar o ciclo reprodutivo.
 |
| Ciclo de vida de um musgo |
 |
| Anterídio |
 |
| Anterídio |
 |
| Anterozoide |
 |
| Cápsula do esporófito de um musgo |
 |
| Polytrichum commune |
 |
| Funaria hygrometrica |
 |
| Sphagnum fallax |
Diferentemente dos musgos, os antóceros possuem um gametófito laminar que lembra uma folha, se desenvolvendo paralelamente ao solo. A sua fixação no solo se dá, assim como no caso dos musgos, através dos rizoides que, no caso dos antóceros, são menores. Assim como os musgos, os antóceros são envoltos por uma camada epidérmica e, em geral, não são muito espessos, o que facilita a troca gasosa.

Quando maduros, os gametófitos dos antóceros desenvolvem as suas estruturas reprodutivas, sendo que, em alguns casos, um mesmo indivíduo pode apresentar tanto a estrutura masculina (isto é, o anterídio), quanto a estrutura feminina (o arquegônio). De maneira similar aos musgos, o anterídio dos antóceros rompe, liberando os anterozoides que, na condição de haver água no ambiente, nadam até o arquegônio feminino, onde se encontra a oosfera. Pela fecundação, se forma o zigoto diploide, que realiza várias mitoses, passando pelas fases de embrião maciço multicelular até diferenciar as suas estruturas esporofíticas. O esporófito dos antóceros se desenvolve quase ereto em relação ao gametófito, geralmente apresentando alguma inclinação. Devido a esta característica, estas espécies são conhecidas, em inglês, por hornworts (horn significando chifre). Na base do esporófito se encontra um tecido meristemático (responsável pelo crescimento nas plantas). As células do esporófito se reproduzem e expandem verticalmente, assim, as células do topo do esporófito são mais velhas do que as da base. Quando as células do esporófito já se encontram maduras, o topo se "rasga", formando uma bifurcação e liberando os esporos ao ar. Quando encontram condições favoráveis, se implantam no substrato e germinam, dando origem ao gametófito.

 |
| Gametófito de um antócero |

 |
| Esporófitos dos antóceros |
Exemplos de antóceros:
c) Hepáticas (liverworts):
 |
| Megaceros aenigmaticus |
 |
| Anthoceros fusiformis |
O gametófito das hepáticas se assemelha ao dos antóceros por ter um corpo do tipo "talo", isto é, um corpo laminar que lembra uma folha ancorado ao solo por meio de rizoides. Como o formato do gametófito lembra um fígado, estas plantas são conhecidas, em inglês, como "liverworts" (liver = fígado em inglês) e hepáticas em português ("hepático" seria qualquer coisa relacionada ao fígado).


Quando as estruturas reprodutivas das hepáticas estão mais amadurecidas, elas já se destacam do restante do talo do gametófito, pois crescem formando uma estrutura que lembra um guarda-chuva, com a parte reprodutiva em si se localizando em seu ápice, sendo que algumas espécies podem apresentar tanto estruturas masculinas quanto femininas. Já amadurecido, o anterídio libera os seus anterozoides, que nadam até o arquegônio feminino e fecundam a oosfera, formando o zigoto diploide, que, por sua vez, cresce em decorrência de várias divisões mitóticas dando origem ao embrião multicelular maciço. Ao contrário do esporófito dos musgos, o esporófito das hepáticas desenvolvem a sua cápsula antes de ocorrer o alongamento da seta (filamento sustenta a cápsula e permite a condução de nutrientes do gametófito para o esporófito). Quando as cápsulas do esporófito se rompem, liberam os esporos para o ambiente, que germinam e formam os gametófitos.











Exemplos de hepáticas:
 |
| Lunularia cruciata |
 |
| Sphaerocarpos texanus |
b) Pteridófitas:
- Primeiros entre as embriófitas a desenvolverem tecidos vasculares para a condução de seiva (xilema e floema)
- Apresentam fase esporófita dominante e gametófito reduzido
- Assim como as briófitas, dependem de água para se reproduzir (gametófito natante) e vivem, portanto, em ambientes úmidos e sombreados, além de não apresentarem sementes e sim esporos
- Já apresentam folhas, caules e raízes verdadeiras
Uma das principais novidades evolutivas apresentadas pelas pteridófitas e que permitiram a expansão destas plantas para além de ambientes úmidos, assim como o aumento do porte destes vegetais é o surgimento de tecidos dedicados à condução de água e nutrientes pelo organismo. Com uma distribuição de sais minerais, aminoácidos e açúcares mais eficiente e rápida, as pteridófitas puderam alcançar tamanhos muito maiores do que as briófitas, cuja transferência destas substâncias era muito mais lenta e limitante, ocorrendo de célula em célula.
Os tecidos condutores, no caso, são o xilema e o floema. O xilema é o responsável por conduzir a chamada seiva bruta (também conhecida por seiva mineral ou seiva xilemática), composta de água e sais minerais, e que parte das raízes (que, nas pteridófitas, têm funções mais importantes que os rizoides das briófitas, pois além de servirem para a fixação do vegetal no substrato terrestre, também absorve os nutrientes encontrados neste meio) em direção às folhas. Já o floema é o canal que conduz a seiva elaborada (seiva orgânica ou, também, seiva floemática), composta de água e produtos da fotossíntese, como açúcares, além de outros nutrientes, como aminoácidos, partindo das folhas e sendo transportados para o restante do organismo. Lembrando que o xilema é formado de células mortas e apresentam paredes celulares mais espessas e repletas de lignina, o que as torna este canal mais rígido que o floema.
Como mencionado, as pteridófitas apresentam raízes, caules e folhas verdadeiras porque estas apresentam sistema vascular. As raízes têm a função de ancorar a planta e absorver água e sais minerais do solo. Através do fenômeno da capilaridade, esta seiva bruta ascende pelo xilema, chegando até as folhas. O caule, nas pteridófitas, geralmente se encontra implantado no subterrâneo, sendo conhecido, então, como rizoma, servindo, também, como estrutura de sustentação para as folhas, que realizam a fotossíntese.





As estruturas reprodutivas do esporófito estão localizadas em suas folhas. As folhas apresentam pequenas estruturas circulares marrons chamadas de soros. Os soros possuem vários esporângios, estruturas que possuem vários esporócitos, que são as células que, por meiose, formam os esporos. O indúsio é a estrutura que envolve os soros.



Os esporos das pteridófitas podem ser de diversos tamanhos e tipos. Espécies que possuem um único tipo de esporo são conhecidas como homosporadas, enquanto as que apresentam mais de um tipo de esporo (esporos maiores, os megásporos, e os menores micrósporos) são as heterosporadas.


 |
| Fronde de pteridófita sem ramificação |
 |
| Fronde de pteridófita ramificada |
{kind=link}
Classificação das pteridófitas: as pteridófitas são classificadas em duas divisões distintas: a divisão Pteridophyta e a divisão Lycopodiophyta.
No clado Pteridophyta (ou também Pterophyta) estão pteridófitas como as samambaias e as avencas. Se caracterizam por suas folhas mais jovens se apresentarem de forma enrolada, também chamada de báculo. Elas também podem ser classificadas como eusporangiadas, em que os esporângios são formados por várias células iniciais, ou leptosporangiadas, quando o esporângio se desenvolve a partir de uma única célula.
Exemplos de pteridófitas:
 |
| Asplenum trichomanes |
 |
As licopodiófitas se distinguem das pteridófitas pois apresentam folhas mais simples. Enquanto as pteridófitas possuem folhas com nervuras ramificadas (as nervuras seriam os "traços" que aparecem na superfície da folha, indicando por onde passam os canais condutores), as licopodiófitas têm folhas com uma única nervura, que não ramifica. Estas folhas mais complexas, características das pteridófitas como as samambaias e os xaxins, são chamadas de megáfilos, enquanto as mais primitivas, típicas das licopodiófitas como as selaginelas, têm o nome de micrófilos.







c) Gimnospermas:
- Primeiras plantas a desenvolver sementes
- O desenvolvimento de grãos de pólen e tubo polínico torna a reprodução das gimnospermas independente da água.
- As sementes das gimnospermas são "nuas", isto é, não são envoltas por um fruto.
- Apresentam esporófito dominante e gametófito reduzido
- Possuem tecido condutor e raízes, folhas e caule verdadeiros
- Possuem esporos de tamanhos diferentes (heterosporia)
Estrutura das gimnospermas:
A principal novidade evolutiva das gimnospermas em relação aos seus antecessores é a semente. A semente é uma estrutura que nasce do desenvolvimento do óvulo (diferente do óvulo dos animais, onde corresponde ao gameta feminino, o óvulo das plantas é uma estrutura complexa formada de tecido do gametófito feminino e do gametófito masculino). As sementes apresentam uma casca resistente que evita a perda de água e, em seu interior, possui reservas de nutrientes para o embrião se alimentar.

 Como as gimnospermas são heterosporadas, os diferentes tipos de esporos estão relacionados a sexos específicos: os esporos menores, ou micrósporos, são produzidos pelos órgãos masculinos enquanto os esporos maiores, ou macrósporos, são produzidos nas estruturas femininas.
Como as gimnospermas são heterosporadas, os diferentes tipos de esporos estão relacionados a sexos específicos: os esporos menores, ou micrósporos, são produzidos pelos órgãos masculinos enquanto os esporos maiores, ou macrósporos, são produzidos nas estruturas femininas.
O estróbilo é outra estrutura reprodutiva importante encontrada nas gimnospermas, sendo responsáveis pela produção dos óvulos nos estróbilos femininos e pela produção e liberação de esporos nos masculinos. Como consequência da diferença de tamanho das estruturas masculinas e femininas, os estróbilos masculinos são menores que os femininos. Os estróbilos também são chamados de "cones" (por isso o nome do grupo "Coniferophyta", um dos mais importantes dentre as gimnospermas) ou "pinhas".





As coníferas correspondem à maior parte das gimnospermas. São, em sua maioria, árvores lenhosas de grande porte e que se caracterizam pela produção de estróbilos (os cones) como estruturas reprodutivas. Alguns exemplos de coníferas são as sequoias, os pinheiros, as araucárias, os lariços, os ciprestes, entre outros.















Exemplos de dicotiledôneas:

Algumas espécies de licopodiófitas são homosporadas, assim como também há espécies heterosporadas, como as do gênero Selaginella.

 |
| Selaginella denticulata |
 |
| Lycopodiella cernua |
Ciclo reprodutivo das pteridófitas:
Ao se implantar no solo, germinam, dando origem ao gametófito haploide, também chamado de prótalo, que possui o formato de um coração. O prótalo se fixa ao solo por meio de rizoides e, ao longo de seu desenvolvimento, forma seus órgãos reprodutivos, sendo que o prótalo dos homosporados é monoico, isto é, apresenta tanto o órgão masculino (o anterídio) quanto o órgão feminino (o arquegônio).

Os anterozoides liberados pelo anterídio nadam até o arquegônio, onde fecundam o gameta feminino, a oosfera, formando um zigoto diploide. Do zigoto se formam o caule e as raízes do esporófito, por meio do qual ele recebe os nutrientes da mãe por um curto período de tempo. Inicialmente, o esporófito jovem se encontra enrolado e, conforme vai envelhecendo, se desenrola na forma de fronde/folha. Enquanto amadurece, o esporófito desenvolve os seus esporângios, até chegar a hora de liberar os seus esporos, reiniciando o ciclo.



- Primeiras plantas a desenvolver sementes
- O desenvolvimento de grãos de pólen e tubo polínico torna a reprodução das gimnospermas independente da água.
- As sementes das gimnospermas são "nuas", isto é, não são envoltas por um fruto.
- Apresentam esporófito dominante e gametófito reduzido
- Possuem tecido condutor e raízes, folhas e caule verdadeiros
- Possuem esporos de tamanhos diferentes (heterosporia)
Estrutura das gimnospermas:
A principal novidade evolutiva das gimnospermas em relação aos seus antecessores é a semente. A semente é uma estrutura que nasce do desenvolvimento do óvulo (diferente do óvulo dos animais, onde corresponde ao gameta feminino, o óvulo das plantas é uma estrutura complexa formada de tecido do gametófito feminino e do gametófito masculino). As sementes apresentam uma casca resistente que evita a perda de água e, em seu interior, possui reservas de nutrientes para o embrião se alimentar.


O estróbilo é outra estrutura reprodutiva importante encontrada nas gimnospermas, sendo responsáveis pela produção dos óvulos nos estróbilos femininos e pela produção e liberação de esporos nos masculinos. Como consequência da diferença de tamanho das estruturas masculinas e femininas, os estróbilos masculinos são menores que os femininos. Os estróbilos também são chamados de "cones" (por isso o nome do grupo "Coniferophyta", um dos mais importantes dentre as gimnospermas) ou "pinhas".

As gimnospermas são, em sua maioria, árvores lenhosas que podem atingir enormes proporções quando comparadas às suas antecessoras briófitas e pteridófitas. A presença de tecido condutor de seiva, além do desenvolvimento da semente e do grão de pólen permitiu que a gimnosperma se tornasse praticamente independente da água para se reproduzir, o que, por sua vez, levou à expansão das plantas cada vez mais em direção aos interiores dos continentes, tendo em vista que não mais precisavam ficar próximas a rios, lagos ou oceanos.
Classificação das gimnospermas: as gimnospermas são divididas em quatro grupos: Cicadophyta (cicas), Coniferophyta (coníferas), Gnetophyta (gnetófitas) e Ginkgophyta (ginkgos).
As cicadófitas possuem folhas cujo tamanho e disposição remetem a uma palmeira. São espécies muito antigas que estiveram presentes na Terra desde a época dos dinossauros.
 |
| Cycas revoluta |






As gnetófitas são arbustos e trepadeiras menores que possuem folhas que são parecidas com as de algumas angiospermas, o que indica uma proximidade evolutiva entre os dois grupos. Um exemplo de gnetófita seriam as plantas do gênero Welwitschia encontradas nas áreas desérticas do sudoeste africano (Angola, Namíbia e África do Sul).


O filo Ginkgophyta é composto por uma única espécie, a Ginkgo biloba, que é considerada um "fóssil vivo" pois existe desde o tempo dos dinossauros (assim como as cicadófitas) e pouco mudou desde então.


Ciclo reprodutivo das gimnospermas:
Como já explicado, a maior novidade que as gimnospermas apresentaram foi a semente, revolucionando os métodos reprodutivos dentre as plantas, que, até então, dependiam totalmente da água para realizar a fecundação. Agora, será discutido o funcionamento destes mecanismos desenvolvidos pelas gimnospermas.
Em alguns indivíduos, se desenvolvem folhas férteis, os megasporofilos, onde se formam os megasporângios. No megasporângio se diferencia um único megasporócito, que está mergulhado em um tecido nutritivo chamado de nucelo e este conjunto é envolvido por uma camada protetora chamada tegumento. O megasporócito passa por meiose, formando quatro células haploides, sendo que três delas se degeneram e a célula restante se diferencia, dando origem ao megásporo.
O megásporo, então, passa por várias mitoses e forma o gametófito feminino, onde se forma o arquegônio, que, por sua vez, forma as oosferas, os gametas femininos. O arquegônio está localizado mais próximo de uma abertura no tegumento, a micrópila, facilitando o contato com o gameta masculino.
As folhas férteis também podem ser do tipo microsporofilo, que forma os microsporângios. No interior dos microsporângios se formam os microsporócitos, que, ao realizar a meiose, formam os micrósporos haploides.
Os micrósporos passam por um processo de mitose, formando quatro células distintas: duas protalares, uma célula do tubo e uma geradora ou germinativa. Este conjunto também é envolvido por uma parede cujo formato favorece o seu transporte pelo vento, sendo chamado, por isso, de "asas". O conjunto formado pelas "asas" e as células originadas da mitose do esporo é o grão de pólen, que é liberado pelos estróbilos para que este chegue até o óvulo e fecunde a oosfera.
O grão de pólen serve como uma nova estratégia reprodutiva, pois seu formato permite ser carregado pelo vento (anemofilia) ou até mesmo por insetos (entomofilia).
Quando o grão de pólen entra em contato com o óvulo, a célula do tubo desenvolve uma estrutura filamentosa que penetra a micrópila e digere o nucelo até chegar ao arquegônio, esta estrutura é o tubo polínico (como algumas gimnospermas desenvolvem o tubo polínico durante o processo reprodutivo, estas são chamadas de sifonógamas). Enquanto o tubo polínico está em crescimento, vai ocorrendo o amadurecimento do gameta masculino. Primeiramente, a célula germinativa passa por mitose, formando uma célula espermatogênica e outra célula estéril. A célula espermatogênica passa por outra mitose, formando dois gametas, sendo que só um deles acaba fecundando a oosfera enquanto o outro se degenera.
Da fecundação da oosfera se forma o zigoto, que se desenvolve formando o embrião. O embrião fica protegido pelo tegumento, que formará a casca da semente, e é nutrido pelo nucelo, o que o torna menos dependente da nutrição da planta-mãe.


d) Angiospermas:
- Primeiras plantas a apresentar flores e frutos
- Assim como as gimnospermas, são espermatófitas, desenvolvendo sementes, e sifonógamas, desenvolvendo grão de pólen e tubo polínico no processo reprodutivo
- Diferentemente das gimnospermas, as suas sementes não estão expostas ao exterior, sendo envolvidas pelo fruto, que surge do desenvolvimento do ovário
- Algumas flores e frutos apresentam diversas formas de atrair animais como insetos e aves para ajudar na polinização e na dispersão das sementes, como cores, cheiros e sabores agradáveis.
- Apresentam esporófito dominante e gametófito reduzido
- Possuem tecidos condutores e estruturas como raízes, caule e folhas verdadeiras
- Apresentam esporos de tamanhos diferentes (heterosporia)
- É o grupo com o maior número de espécies do reino vegetal
Classificação das angiospermas:
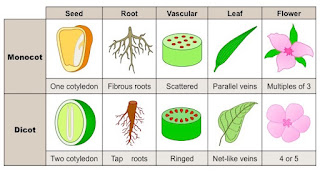
As angiospermas são classificadas em dois grupos: as dicotiledôneas e as monocotiledôneas, divisão esta que se dá pelo número de cotilédones, que são folhas que cumprem função de reserva nutritiva no embrião, que se desenvolvem em cada planta (as monocotiledôneas desenvolvem um único cotilédone e as dicotiledôneas desenvolvem dois cotilédones). Diversos traços distintos nos permitem identificar a qual grupo uma angiosperma pertence, dentre elas:
- A nervura das folhas (paralelas nas monocotiledôneas e ramificadas nas dicotiledôneas).

Analisando a folha do bambu, percebemos que as nervuras de sua folha são paralelas em relação ao eixo principal desta estrutura, logo, podemos concluir que o bambu pertence ao grupo das monocotiledôneas.

A folha de laranjeira apresenta nervuras ramificadas, logo, esta planta pertence ao grupo das dicotiledôneas.
- Vasos condutores dispostos em anéis (no caso das dicotiledôneas) ou espalhados (no caso das monocotiledôneas), o que pode ser percebido na formação, ou não, de anéis de crescimento. Outro fator que influencia neste diferencial, é que as monocotiledôneas não apresentam o câmbio, tecido que praticamente comanda o aumento da espessura da planta, também chamado de crescimento secundário, apresentando somente, então, o crescimento primário, em que ocorre o aumento do comprimento da planta. Ao longo de seu crescimento secundário, as dicotiledôneas produzem anéis que refletem a variação das estações e as mudanças do tempo, podendo ser utilizadas para descobrir a idade das árvores.
 |
| O coqueiro, por ser uma monocotiledônea, não apresenta anéis de crescimento |

- As flores das monocotiledôneas são formadas por um número de partes múltiplo de 3, enquanto as flores das dicotiledôneas são formadas por partes múltiplas de 4 ou 5.


Exemplos de monocotiledôneas:
 |
| Gramíneas |
 |
| Coqueiros |
 |
| Orquídeas |
 |
| Tulipas |
 |
| Bananeira |
Exemplos de dicotiledôneas:
 |
| Girassol |
 |
| Rosas |
 |
| Cafeeiro |
 |
| Macieira |
 |
| Feijoeiro |
As flores são uma novidade para as plantas, cumprindo um papel parecido com o dos estróbilos nas gimnospermas, sendo compostas por folhas especiais onde são produzidas as estruturas reprodutivas masculinas e femininas.
A flor é contida por um ramo chamado de pedicelo. Em sua ponta está o receptáculo floral, que contém as folhas especiais da flor, os verticilos florais. Uma flor é considerada completa quando apresenta todos os quatro verticilos florais: cálice, corola, androceu e gineceu.
O cálice e a corola formam o perianto, e as suas folhas modificadas são, respectivamente, as sépalas e as pétalas. As sépalas são menores e de cor verde enquanto as pétalas são maiores e coloridas, ajudando na atração de insetos, aves, humanos, etc.
O androceu e o gineceu são os conjuntos de estruturas reprodutivas masculinas e femininas, respectivamente. As folhas do androceu são os estames, que são compostos pela antera e a sua estrutura filamentosa de sustentação, o filete. Na antera se encontram os microsporângios, que produzem os microsporócitos, que, por sua vez, dão origem aos micrósporos, chegando finalmente aos grãos de pólen. As folhas do gineceu são os pistilos, que são compostos por estigma, estilete e ovário. O estigma é a estrutura com a qual os grãos de pólen fazem contato. O estilete é um tubo pelo qual o tubo polínico cresce até chegar ao ovário, onde se encontram os óvulos.


Durante o amadurecimento da flor, surgem protuberâncias na parede do ovário, que dão origem aos óvulos. Em cada um destes óvulos uma célula acaba se tornando predominante, sendo a precursora do megasporócito. Ao redor do megasporócito se desenvolvem as células do nucelo, o tecido nutritivo, e as do tegumento, a camada externa protetora, em que se forma uma pequena abertura, a micrópila, por onde penetrará o tubo polínico e entrarão os gametas masculinos.
O megasporócito eventualmente passa por meiose, produzindo quatro células haploides (n), das quais três se degeneram e a última célula restante se diferencia, formando o megásporo. Este megásporo passa por várias mitoses, formando o megagametófito ou gametófito feminino. Destas mitoses se formam oito núcleos. Oito destes núcleos migram mais próximas à micrópila, enquanto os demais migram para o polo oposto. Um núcleo de cada um destes conjuntos se deslocam para o centro do megásporo, aonde se fundem formando uma célula diploide (2n). Se forma no megásporo, então, uma configuração característica: três células haploides em um polo próximo à micrópila (duas destas são chamadas de sinérgides enquanto a central é a oosfera, o gameta feminino em si) e outras três células haploides no polo oposto, as antípodas, com o restante do espaço sendo ocupado pela célula central diploide composta pelos dois núcleos polares.
Enquanto isso, nos microsporângios das anteras são produzidos os microsporócitos, que passam por meiose formando os micrósporos. Os micrósporos por sua vez passam por mitose, levando à formação do grão de pólen (nas angiospermas, diferentemente das gimnospermas, são compostas por uma célula do tubo e uma célula germinativa somente, esta última se divide formando os dois gametas masculinos, os núcleos espermáticos).
Quando a antera se encontra amadurecida, libera os grãos de pólen, que são carregados pela ação do vento (anemofilia), insetos como as abelhas (entomofilia), entre outros agentes, até entrarem em contato com o gineceu de uma flor. Ao fazerem contato com o estigma, o grão de pólen absorve a solução açucarada produzida pelo estigma e germina, formando o tubo polínico, que vai penetrando pelo estilete em busca do óvulo. Ao chegar no óvulo, um dos núcleo espermáticos fecunda a oosfera, formando o zigoto diploide (2n), que dará origem ao embrião, enquanto o outro núcleo espermático se funde aos núcleos polares, formando a endosperma triploide (3n), que tem função principalmente nutritiva na semente. Como nas angiospermas ocorrem duas fecundações diferentes: a da oosfera e a dos núcleos polares, este fenômeno é chamado de dupla fecundação.


Durante o desenvolvimento do embrião, se formam partes distintas: a radícula, que é precursora das raízes; o hipocótilo, que corresponderá à parte inferior do caule, e o epicótilo, que corresponderá à parte superior do caule. Estas duas últimas partes são delimitadas pelos cotilédones, que as separam.




O fruto resulta do desenvolvimento do ovário das angiospermas, protegendo as sementes em seus interiores e ajudando na dispersão destas através dos animais, por exemplo, que ingerem a fruta e liberam as sementes durante a defecação. Algumas estruturas que costumamos chamar de "frutos" são, na verdade, pseudofrutos ou frutos acessórios, pois resultam do desenvolvimento de uma estrutura anexa ou associada ao fruto, como é o caso do caju. Outros frutos, os chamados frutos partenocárpicos, também se desenvolvem sem a ocorrência de fecundação, assim, não apresentam sementes, como é o caso da banana (a banana, antigamente, era um fruto menor e com uma grande quantidade de sementes grandes, porém, com a sua domesticação pelos humanos, suas sementes se tornaram cada vez menores, com uma parte comestível maior).


Com o desenvolvimento do embrião, a parede do ovário se diferencia, formando o pericarpo. O pericarpo é dividido em três partes, de dentro para fora: endocarpo, mesocarpo e exocarpo. Os frutos que apresentam pericarpo mais desenvolvido são chamados de frutos carnosos (como a laranja, a uva, o pêssego, entre outros) enquanto os que apresentam pericarpo subdesenvolvido são os frutos secos, como a vagem do feijão.
Os frutos podem ser classificados em:
- Frutos simples: são os frutos verdadeiros que se formam do desenvolvimento das estruturas de uma úncia flor. Podem ser classificados em carnosos, quando apresentam pericarpo desenvolvido com mesocarpo suculento, ou secos, com pericarpo subdesenvolvido.


- Frutos agregados: formado por vários ovários amadurecidos, pode se dizer que se tratam de vários frutos simples contidos em uma outra estrutura mais desenvolvida (como no caso do morango, em que os frutos verdadeiros são os "pontinhos amarelos" enquanto a parte que costumamos chamar de "fruto" é o receptáculo floral desenvolvido).


Anatomia e fisiologia vegetal:
Nesta seção, alguns conceitos relativos às estruturas das plantas e suas respectativas funções e funcionamentos ainda não explorados serão tratados.
As plantas se assemelham com os animais no aspecto de que ambos os grupos apresentam tecidos especificados que cumprem determinadas funções. Nas plantas, estes tecidos podem ser classificados nos seguintes grupos: tecidos de revestimento, tecidos de condução, tecidos de preenchimento, tecidos de sustentação e os tecidos meristemáticos.
Tecidos de revestimento:
Os tecidos de revestimento são a epiderme e a periderme, que, basicamente, revestem a planta cumprindo uma variedade de funções, desde a proteção até as trocas gasosas. Por serem de revestimento, são os tecidos que se encontram nas porções mais externas da planta.
A epiderme é constituída de células vivas encontrada nas porções externas das folhas, frutos e flores, além dos caules e raízes das plantas com crescimento primário (ou seja, em comprimento). As células da epiderme geralmente secretam uma substância cerosa, a cutina, que forma a cutícula, estrutura que ajuda a evitar a perda de água para o ambiente, além de atuar na impermeabilização e servir como mais uma forma de impedir a entrada de agentes invasores na planta.


Como já mencionado, os estômatos são as estruturas que as plantas apresentam para realizar as trocas gasosas. O controle da abertura dos estômatos é feito pelas chamadas células-guarda, que se abrem quando absorvem muita água, permitindo a entrada do gás carbônico necessário para a fotossíntese, além da saída de gás oxigênio e água por transpiração. Em condições mais adversas, como tempos mais quentes, as células-guarda são orientadas a não ficarem abertas por muito tempo para evitar a perda excessiva de água, assim a epiderme atua tanto nas trocas gasosas quanto no processo de transpiração.
Algumas plantas também apresentam tricomas, acúleos, espinhos e hidatódios como estruturas anexas da epiderme. Estes apêndices podem ter diversas funções, dentre elas estão: absorção de água e umidade do ar (como nos espinhos dos cactos), proteção (no caso das estruturas enrijecidas e pontudas que não possuem tecido vascular, os acúleos, que atuam como espinhos, ajudando a afugentar animais, protegendo a planta. Um exemplo de planta onde ocorrem os acúleos são as rosas), secreção de substâncias, desde as ceras, para impermeabilizar a planta, eliminação de componentes tóxicos em excesso, substâncias que repelem ou capturam insetos (como nas plantas carnívoras), e eliminação de água em excesso através do processo de gutação no caso dos hidatódios.




A periderme ocorre nas plantas que apresentam crescimento secundário (isto é, em que a espessura da planta aumenta) e se caracteriza por ser mais complexa que a epiderme, tendo em vista que é composta por mais de um tipo de tecido: o súber, o felogênio e a feloderme.
Este tecido tem sua formação iniciada com base no felogênio, que se trata de um tecido meristemático, ou seja, que comanda o processo de divisão e diferenciação celular na planta. Para a porção interior da planta, se forma a feloderme, constituída de células parenquimatosas que atuam no preenchimento e acumulação de reservas, já para a porção exterior, se forma o súber, que é composto por células bem unidas e compactadas que produzem uma substância chamada suberina. A suberina, assim como a cutina, é de caráter lipídico e altamente hidrofóbica, garantindo a impermeabilização da planta. Contudo, conforme as células do súber amadurecem, esta substância se acumula, impedindo até mesmo a chegada de nutrientes da planta até ela, resultando em sua morte, deixando somente a sua parede celular suberificada. Esta camada de células mortas da periderme corresponde à casca da árvore.
Se a periderme apresenta como camada mais externa um conjunto de células mortas altamente compactadas, como são feitas as trocas gasosas? Para a realização de tal função tão vital, o felogênio forma aberturas nos caules e nas raízes, as lenticelas, que permitem a entrada e saída de gases na planta.


 |
| Lenticelas |

Tecidos de condução:
Os tecidos de condução (ou tecidos vasculares), se assemelham, em função, ao sistema circulatório dos animais, pois transportam substâncias e nutrientes necessárias para a sobrevivência das células. Estes tecidos são o xilema e o floema.
O xilema conduz a seiva bruta, que é composta basicamente por água e sais minerais e é conduzido das raízes até as folhas. Este tecido é constituído por células traqueais, fibras e células do parênquima, que atuam como armazenamentos de substâncias e fibras. As células traqueais, os traqueideos e os elementos do vaso, são ambas células alongadas e tubulares por onde passa a seiva bruta ou xilemática. Estas células também apresentam uma parede celular secundária, esta que armazena uma substância chamada lignina, que confere rigidez e impermeabilidade a estas células (por causa desta substância, o xilema também é chamado de lenho). A lignina é um dos principais componentes da madeira, sendo, portanto, importante para a sustentação das plantas lenhosas. O acúmulo de lignina nestas células, ao longo de seu amadurecimento, culmina na morte destas células, que se desfazem, então, de seus componentes internos (núcleo, citoplasma, organelas, etc.), deixando apenas as suas paredes celulares de celulose e lignina, atuando, basicamente, como tubos por onde passa a seiva. As células traqueais se comunicam por meio de aberturas em suas paredes celulares ou em pontos onde a parede secundária é menos espessa, permitindo a passagem de água de célula a célula.


O floema conduz a seiva elaborada, que carrega compostos orgânicos como aminoácidos, proteínas, RNA, hormônios, lipídios, entre outros, além dos produtos da fotossíntese, que são levados das folhas até as demais células do corpo da planta. O floema é composto por dois tipos distintos de células: os elementos crivados e as células companheiras. Os elementos crivados se chamam assim pois apresentam uma série de poros em suas extremidades, sendo que é por meio destes que se faz o transporte da seiva de célula em célula. Ao contrário das células do lenho, as células do floema (ou também, liber) não se encontram mortas em sua maturidade, porém, não apresentam núcleo, ribossomos e outras estruturas celulares, portanto, dependem do apoio das células companheiras, que os fornecem substâncias e nutrientes.


A localização dos tecidos condutores entre eles varia dependendo da estrutura da planta. No caule, o xilema se encontra mais interior do que o floema, sendo que os dois frequentemente estão dispostos próximos ao outro, formando o feixe vascular. Nas folhas, o vaso condutor que se encontra mais próximo em relação ao centro do caule, neste caso, o xilema, se encontra na posição superior, enquanto o floema está na posição inferior. Nas raízes, o xilema está disposto em uma forma de cruz ou "X", com o floema preenchendo o restante do feixe vascular, (também chamado de estela nas raízes).


O funcionamento do sistema condutor de seiva das plantas é intrigante pois, diferentemente dos animais, as plantas não apresentam uma forma de "bombear" este líquido, portanto, precisam utilizar de outros meios para garantir o transporte de água e nutrientes ao redor de seu corpo enfrentando a força da gravidade:
As raízes da planta apresentam uma maior concentração de soluto nas suas células do que o solo, portanto, a água do solo entra nas raízes por osmose (meio menos concentrado -> mais concentrado). A entrada de água resulta em um aumento de pressão nas raízes fazendo com que a água seja empurrada para cima.

Nos caules mais estreitos, a água pode ascender através do fenômeno da capilaridade ou ação capilar, em que, por meio das forças de coesão (o que causa a tensão superficial, em que a superfície da água se torna, essencialmente, uma película fina) e adesão (em que as moléculas de água se atraem às paredes do recipiente e acabam se aderindo a elas) a água sobe pelo caule.

A porção final do processo de subida da água pelo xilema ocorre através da transpiração. A evaporação que ocorre através da abertura dos estômatos faz com que a pressão nas folhas diminua. Assim, a água, que se direciona de ambientes de maior pressão para os de menor pressão, acaba sendo "puxada" até as folhas.

No floema, o transporte de seiva se dá por meio da diferença da concentração de soluto entre os "carregadores" e os "descarregadores" de açúcares da planta. A produção de açúcares ocorre nas folhas através da fotossíntese, portanto, estas são consideradas fontes ou "carregadoras" pois depositam estes açúcares (além de outros nutrientes) no floema, que os levam até as raízes, que são os "descarregadores" ou "drenos" de açúcares. Em algumas situações, o fluxo do floema (que, normalmente, é de cima - partindo das folhas - para baixo - nas raízes) pode ser invertido, como no caso do ápice da planta em que, por causa do crescimento, há uma demanda maior por nutrientes que, quando consumidos, provocam uma diferença entre a pressão de soluto do ápice e das folhas, tornando o primeiro um "dreno", fazendo com que a seiva seja deslocada para cima. O processo de deslocamento da seiva elaborada é chamado de "translocação", sendo que este pode se dar através das rotas simplástica ou apoplástica. A diferença entre estes dois modos é que, basicamente, na rota simplástica os nutrientes passam de célula em célula através de seus citoplasmas enquanto na apoplástica este movimento se pelas paredes celulares.


Tecidos de preenchimento:
Os tecidos de preenchimento correspondem à grande variedade de tipos de parênquimas existentes na planta. As células do parênquima se caracterizam por uma enorme capacidade de divisão e diferenciação, assim, dependendo da situação, podem se converter em determinados tipos de célula e integrar os órgãos da planta. Além disso, as principais funções desempenhadas por estas células são as de formar a maior parte da massa da planta (por isso o parênquima é chamado de tecido de preenchimento) além de armazenar substâncias que são importantes para o seu funcionamento.
As células do parênquima encontradas nas folhas formam o mesofilo, camada que se encontra abaixo da epiderme e é especializada para realizar a fotossíntese (por isso, o parênquima das folhas também é chamado de clorênquima). As células do clorênquima se dividem em dois tipos: as células paliçádicas e as células esponjosas ou lacunosas. As paliçádicas são mais compactas e se encontram nas porções mais externas da folha, assim, captam mais raios luminosos e, portanto, têm atividade fotossintética mais intensa. Já as células esponjosas estão mais internas na folha e apresentam lacunas maiores entre elas, permitindo a circulação de água e gases dentro da folha.


Encontrado em diversas partes da planta, o parênquima de reserva sintetiza e armazena diversas substâncias em seus vacúolos e em seus citoplasmas. Este tipo de parênquima pode ser dividido em três subtipos: o parênquima amilífero, o aquífero e o aerífero (ou aerênquima). O amilífero armazena o amido, um polissacarídeo formado a partir de várias moléculas de glicose que serve como principal reserva de energia nas plantas, o aquífero armazena água e as células do aerífero apresentam grandes espaços intercelulares, permitindo a circulação de gás por estes espaços, o que, em alguns casos, permite que a planta flutue na água.



Tecidos de sustentação:
Os tecidos de sustentação, o clorênquima e o esclerênquima, conferem resistência e sustentação às suas partes da planta.
O clorênquima é composto por células vivas com paredes celulares espessadas por celulose de modo irregular. As células do esclerênquima, por sua vez, são células mortas desprovidas de protoplasma, que apresentam uma parede celular secundária composta de celulose e lignina distribuídas igualmente e, além disso, apresentam pouco espaço intercelular, portanto, são muito compactas. Como o esclerênquima apresenta uma segunda parede celular que, ainda por cima, possui lignina em sua composição, as suas células são muito mais rígidas que as do clorênquima, estas que se caracterizam por apresentar certa flexibilidade, o que permite o alongamento e o crescimento das estruturas da planta. O clorênquima, também, está presente na folha localizada próxima a epiderme do órgão e, como se trata de um conjunto de células vivas, estas também realizam fotossíntese. O esclerênquima está, também, associado à casca da árvore.

Tecido meristemático
Os tecidos meristemáticos são os responsáveis por provocar o crescimento e desenvolvimento dos órgãos da planta. Esta capacidade de suas células provém do fato de que todas elas são células indiferenciadas (ou seja, que não possuem uma função definida além de se multiplicarem) que podem se diferenciar em qualquer tipo de célula vegetal, formando tecidos diferentes.
Os tecidos meristemáticos, portanto, se encontram nas partes da planta em que há um constante crescimento, como nas pontas do caule e das raízes, por exemplo.
Este tecido pode ser classificado em diversos tipos dependendo de quais tipos de plantas aparecem e em qual fase do desenvolvimento destas ele é formado:
Meristemas primários: se formam do desenvolvimento embrionário da planta e são responsáveis pelo crescimento primário da planta, ou seja, em que ocorre um aumento do corpo no sentido de seu comprimento. Este processo é realizado pelos meristemas apicais do caule e da raiz. O primeiro promove a divisão das células para a formação das folhas, flores e frutos. O segundo promove a divisão das células na raiz levando ao seu crescimento. Como o crescimento das raízes se dá no subsolo, estas enfrentam o intenso atrito das partículas do solo, assim, para minimizar o seu desgaste, elas são envolvidas, em suas pontas, por uma espécie de "capuz" de células, a coifa, que protege as células meristemáticas da raiz, garantindo o seu crescimento. Desta maneira, o meristema apical do caule se diferencia do da raiz, tendo em vista que o primeiro não apresenta alguma forma de proteção já que não sofre com barreiras sólidas por estar crescendo contra o ar.


Os meristemas apicais formam os meristemas primários: a protoderme, localizada nas partes mais externas do caule, folha e raiz que origina a epiderme; o procâmbio, que forma os xilema e floema primários, e o meristema fundamental, que dá origem aos tecidos de preenchimento e sustentação: parênquima, colênquima e esclerênquima.


Meristemas secundários: se desenvolvem nas plantas com crescimento secundário (em espessura). Os dois tecidos meristemáticos deste tipo são os câmbios vascular e cortical (ou felogênio). O câmbio vascular se instala entre o xilema e o floema primários produzindo os xilema e floema secundários (ao todo, os tecidos vasculares se dispõem de dentro para fora, nesta ordem: xilema primário, xilema secundário, floema secundário e floema primário). O felogênio se situa anterior à epiderme e forma a feloderme (interior) e o súber (exterior), constituindo a periderme.


Substâncias e nutrientes importantes para o crescimento e funcionamento das plantas:
Micro e macronutrientes:
Além de água, luz e ar, para que uma planta cresça saudável é preciso garantir o fornecimento de uma série de elementos inorgânicos que desempenham diversas tarefas no organismo. Estas substâncias podem ser classificadas de acordo com a quantidade necessária para um melhor funcionamento e desenvolvimento da planta: os macronutrientes são aqueles que devem ser providos em grandes quantidades (macro = grande, extenso), enquanto para os micronutrientes basta uma quantidade menor (micro = pequeno, menor).
Macronutrientes estruturais (H, C e O): Alguns macronutrientes já foram, de certa forma, discutidos anteriormente, estes sendo o carbono (C), o hidrogênio (H) e o oxigênio (O). Estes três elementos são principalmente observados nos processos de nutrição e metabolismo da planta, em que são rearranjados para formar açúcares e gases que, em seguida, são usados para gerar a energia necessária para o crescimento das estruturas do vegetal. Têm papel importante, também, na composição de biomoléculas que formam o grosso das células e tecidos do organismo, portanto, são conhecidos como os macronutrientes estruturais da planta.
Macronutrientes primários (N, P e K): Nitrogênio (N), fósforo (P) e potássio (K) são os chamados macronutrientes primários, pois são demandados a volumes maiores que os secundários.
O nitrogênio é um dos mais importantes dentre eles, pois participa da composição de aminoácidos e, consequentemente, das proteínas também. Por isso, o nitrogênio também cumpre função fundamental na estrutura dos vegetais. Esse elemento também compõe os cloroplastos da planta, outro fator que o torna ainda mais importante pois estas estruturas são as responsáveis pela absorção da luz necessária para a fotossíntese. Uma deficiência de nitrogênio na planta pode ser extremamente danosa, pois, com uma menor quantidade deste nutriente, menos cloroplastos podem ser produzidos e, assim, menos intensa é a fotossíntese, tornando o seu desenvolvimento ainda mais reduzido. Como o nitrogênio também faz parte das proteínas, a sua deficiência pode fazer a planta mais frágil e menos resistente. Ressalta-se aqui o papel das bactérias na fixação de nitrogênio, que enriquece o solo com este nutriente, garantindo o fornecimento desta substância à planta.


 |
| Em decorrência de um menor número de cloroplastos, as plantas com deficiência de nitrogênio perdem a sua coloração esverdeada e começam a "amarelar". O potássio tem função importante para controlar a abertura dos estômatos (são os íons que provocam o aumento da turgidez das células-guarda) e, portanto, também regula as trocas gasosas e a perda de água. É importante, também, para a realização da fotossíntese, ativação de enzimas, resistência ao frio e pragas e para a formação de raízes, folhas e frutos. A deficiência deste macronutriente pode fazer a planta mais sujeita ao desenvolvimento de doenças e ser afetada mais facilmente pelo frio. Em menor concentração, provoca o desregulamento da perda de água e das trocas gasosas, alterando a fotossíntese e, consequentemente, o crescimento da planta. Um dos principais sintomas da deficiência de potássio nas plantas é a ocorrência da clorose, em que a cor da folha passa de um tom esverdeado para um mais amarelado, neste caso iniciando pelas bordas da folha. Quando esta mudança de cor avança de modo a atingir as nervuras da folha, esta pode acabar necrosando (ficando com uma cor castanha ou "amarronzada"), se curvar para baixo e até se desprender da planta e cair. |



O fósforo participa da composição do material genético, fazendo parte do grupo fosfato que compõe os ácidos nucleicos, portanto, é importante para que a divisão celular das plantas seja concretizada com êxito, garantindo o seu crescimento. Além dos ácidos nucleicos, o fósforo faz parte do ATP (adenosina trifosfato), molécula em cujas ligações fica armazenada a energia produzida pela planta, portanto, o fósforo também é importante para a conversão e transferência de energia. A deficiência de fósforo pode ser percebida pelo crescimento reduzido das folhas e também pela formação de manchas escuras de tom púrpura nas folhas mais velhas em decorrência da síntese de um pigmento chamado antocianina, produzido como consequência dos açúcares que ficam acumulados nos tecidos.


Macronutrientes secundários (Ca, Mg e S): os macronutrientes secundários são nutrientes que a planta precisa a uma menor quantidade que os macronutrientes primários porém, a uma quantidade maior que os micronutrientes. Estes são o cálcio (Ca), magnésio (Mg) e enxofre (S).
O cálcio participa no crescimento das raízes, germinação do grão de pólen, transporte de nutrientes, e na síntese e ativação de enzimas. Faz parte, também, da parede celular e tem papel importante para a permeabilidade seletiva da membrana das plantas. Nota-se, além disso, a importância do cálcio em reduzir a acidez do solo, facilitando o cultivo em solos mais ácidos (como é o caso do Brasil). A deficiência deste mineral pode, portanto, diminuir o crescimento das raízes (minimizando a absorção de nutrientes, causando um menor crescimento da planta) e dificultar a germinação do pólen. Se observa o enrolamento das folhas, além de casos de clorose.


A importância do magnésio se deve ao fato que este mineral compõe a substância clorofila, pigmento responsável pela absorção de luz dos cloroplastos, portanto, o magnésio é fundamental para a realização da fotossíntese (sem magnésio, não há clorofila, sem clorofila não há cloroplastos e sem cloroplastos não há fotossíntese). A deficiência de magnésio se torna notável pela cloração das plantas, passando por tons amarelos que podem evoluir a manchas avermelhadas que podem acabar necrosando. Com uma redução da clorofila, o crescimento da planta é afetado e, assim, folhas, flores e frutos ficam menores e menos desenvolvidos.


O enxofre participa da formação de aminoácidos, proteínas e enzimas, além de auxiliar na resistência a parasitas e ao frio. A deficiência de enxofre causa uma redução do crescimento das raízes, folhas, flores e frutos, que ficam menores e menos resistentes (bastante parecida com a deficiência de nitrogênio). A clorose se dá principalmente nas folhas mais novas


Micronutrientes: nutrientes que a planta necessita nos menores volumes, estes são: ferro (Fe), manganês (Mn), boro (B), cobre (Cu), zinco (Zn), cloro (Cl) e molibdênio (Mo).
O ferro atua como catalisador da síntese de clorofila e como transportador de oxigênio (é necessário, portanto, para a fotossíntese e para a respiração celular), além de participar da formação e das funções das enzimas. A falta de ferro nas plantas pode causar a deficiência da fotossíntese, observável nas folhas mais novas, que ficam amareladas e frágeis.


O manganês tem várias funções parecidas com a do ferro, participando do processo de síntese de enzimas e estruturas que participam da fotossíntese, além da produção de aminoácidos. Por isso, os sintomas de carência de manganês são bastante parecidos com os do ferro (folhas reduzidas e cor amarelada entre as nervuras da folha)

O boro participa da metabolização dos ácidos nucleicos que compõem o RNA e o DNA, portanto, é um micronutriente importante para a divisão celular, além de ser necessário para o transporte de açúcares e para a formação das paredes celulares de forma que a sua carência pode afetar o crescimento da planta e as suas estruturas (folhas, caule, raízes, frutos, flores, sementes, etc.). Nota-se o desbotamento da coloração das pontas das folhas, que secam e começam a se "enrolar".


O cobre é essencial à fotossíntese e à produção da clorofila. Atua, também, na produção das paredes celulares e das sementes, além de auxiliar na fixação do nitrogênio no solo. Sua deficiência causa o murchamento das folhas, que formam pequenas manchas de coloração mais escura em suas pontas.

O zinco atua como ativador de enzimas e importante transportador de material genético, sendo, então, fundamental para o crescimento da planta. A deficiência faz com que as folhas fiquem com o seu crescimento impactado e apresentem cloração entre as suas nervuras.


O cloro é importante para a fotossíntese, mais especificamente durante o processo de separação da água em hidrogênio e oxigênio. Além disso, também participa da regulação da osmose das células (torna os meios hiper ou hipotônicos). A deficiência do cloro geralmente é rara, porém, quando ocorre, provoca murchamento, clorose e necrose das folhas.

Por último, o molibdênio participa na fixação do nitrogênio no solo, auxiliando na síntese de aminoácidos e proteínas. A deficiência de molibdênio, portanto, pode provocar a deficiência de nitrogênio, causando seus respectivos sintomas (estruturas frágeis, clorose das folhas, crescimento afetado, etc.)




Hormônios vegetais:
Por fim, temos os hormônios vegetais, substâncias químicas que regulam o processo de funcionamento e crescimento das estruturas das plantas.
Auxinas:
Um dos principais hormônios vegetais (ou fitormônios), atuam especialmente no crescimento da planta em relação a uma série de fatores. A auxina mais comum é o AIA, o ácido indolilacético, que é sintetizado nas estruturas mais jovens da planta (folhas e porção apical do caule).

O AIA atua no processo de dominância apical. Durante o crescimento longitudinal, isto é, para cima, o hormônio inibe os tecidos que provocam a germinação lateral. Posteriormente, quando a gema apical, responsável pela síntese do AIA, é eliminada, as gemas laterais se tornam ativadas e provocam a expansão da planta predominantemente "para os lados", gerando vários galhos, folhas e frutos, formando a copa da árvore).

Outro papel importante das auxinas é o crescimento da planta em direção à luz (fototropismo). Quando um lado da planta recebe mais luz do que o outro lado, as auxinas migram para o lado menos iluminado e provocam a distensão ou alongamento destas células, fazendo com que o corpo da planta se curve na direção da fonte de luz assim garantindo uma melhor absorção de luz.


As auxinas atuam, de forma geral, no crescimento de frutos, folhas, raízes e caules, podendo provocar fenômenos como a partenocarpia, em que um fruto se desenvolve sem necessidade de fecundação (exemplo: bananas)

Conforme os frutos e folhas vão amadurecendo, as suas sínteses de auxina vão diminuindo enquanto a produção de outro fitormônio, o etileno, vai aumentando e inibindo o transporte de auxina no local. Como consequência disso, o etileno acaba provocando a dissolução da parede celular das células do pecíolo (segmento que liga a folha e o fruto ao seu ramo), o enfraquecendo e facilitando a queda da folha ou do fruto. As auxinas, assim como o etileno, então, influenciam a queda ou abscisão das folhas e das frutas das plantas.


As auxinas também são responsáveis pelo geotropismo ou gravitropismo, fenômeno em que o caule apresenta crescimento contrário à força da gravidade e a raiz cresce na direção desta força.


Giberelinas:
Assim como as auxinas, as giberelinas são importantes para o crescimento das plantas e suas respectivas estruturas através do alongamento das células e da divisão celular. Um aspecto que destaca as giberelinas é o seu papel em "ativar" estruturas "dormentes". A "dormência", nas plantas, significa um estado em que uma estrutura germinativa (como gemas apicais e laterais e as sementes) tem o seu crescimento inibido. As giberelinas portanto, tiram as gemas e as sementes de seus estados de dormência e incentivam os seus desenvolvimentos. Desta forma, estes hormônios são fundamentais para a germinação de novas plantas e também podem ser aplicados em plantas "anãs" para que um crescimento normal seja estimulado.

Citocininas:
São hormônios sintetizados no ápice das raízes e são enviados "planta acima" através do xilema, (além de serem produzidos em frutos e folhas jovens), atuando no crescimento do organismo incentivando as divisões celulares. Assim como as auxinas, as citocininas participam do controle da dominância apical. As gemas laterias são inibidas pelas auxinas, e, quando a gema apical cessa suas funções, as citocininas são transportadas pelo xilema e ativando as gemas laterais, provocando o crescimento lateral da planta. Estes hormônios também são importantes para retardar o envelhecimento da planta (senescência), portanto, podem ser utilizados para conservá-las.


Etileno:
Como mencionado, o etileno participa do processo de queda de folhas e frutos. O aumento da síntese de etileno acaba provocando a ação de enzimas que digerem as paredes celulares, tornando os ramos que ligam as folhas e os frutos à planta mais frágeis, fazendo com que estas se desprendam ou caiam mais facilmente. O etileno também está associado ao amadurecimento das frutas, provocando a mudança de cor, o amolecimento da fruta e o acúmulo de açúcares. Por causa disso, o etileno pode ser empregado na fruticultura, acelerando o amadurecimento das frutas, ou evitado, a fim de que as frutas não amadureçam e deteriorem antes da comercialização.




Ácido abscísico
O ácido abscísico (ABA) é um hormônio vegetal que inibe o crescimento das plantas, podendo induzir o estado de dormência nas estruturas germinativas, como as sementes. Esta função pode ajudar a planta a sobreviver durante períodos de condições infavoráveis (temperaturas muito altas ou muito baixas, secas, etc.). Também está relacionado à abertura e fechamento dos estômatos das células, evitando a perda de água, o que ajuda a planta a resistir tempos com poucas chuvas. Pode-se considerar, então, o ácido abscísico como um hormônio principalmente de defesa da planta, pois auxilia na sua conservação em períodos mais extremos.



https://www.britannica.com/biography/August-Wilhelm-Eichler
http://segundocientista.blogspot.com/2015/09/botanica-scientia-amabilis.html
https://www.sobiologia.com.br/conteudos/bioquimica/bioquimica10.php
https://www.infoescola.com/citologia/tilacoide/
https://www.nature.com/scitable/topicpage/plant-cells-chloroplasts-and-cell-walls-14053956/
https://courses.lumenlearning.com/wm-biology1/chapter/reading-unique-features-of-plant-cells/
https://ssec.si.edu/stemvisions-blog/what-photosynthesis
https://www.biology-pages.info/G/GasExchange.html
https://www.sparknotes.com/biology/plants/plantstructures/section4/
http://www.webexhibits.org/causesofcolor/7A.html
https://www.sobiologia.com.br/conteudos/bioquimica/bioquimica14.php
https://www.sobiologia.com.br/conteudos/bioquimica/bioquimica15.php
http://educacao.globo.com/biologia/assunto/fisiologia-celular/fotossintese.html
https://www.sobiologia.com.br/conteudos/bioquimica/bioquimica16.php
https://www.peoi.org/Courses/Coursesen/bot/temp/bot16t91.html
https://sciencing.com/how-do-nonvascular-plants-get-water-nutrients-13428005.html
http://www3.botany.ubc.ca/bryophyte/mossintro.html
http://www.criptogamas.ib.ufu.br/node/546
https://courses.lumenlearning.com/wm-biology2/chapter/bryophytes/
http://universe-review.ca/R10-34-anatomy202.htm
https://bio.libretexts.org/Bookshelves/Introductory_and_General_Biology/Book%3A_General_Biology_(Boundless)/25%3A_Seedless_Plants/25.3%3A_Bryophytes/25.3B%3A_Liverworts_and_Hornworts
http://plantphys.info/plant_biology/lechtml/hornwort/
https://cronodon.com/BioTech/liverworts.html
https://www.britannica.com/topic/list-of-mosses-2075729
https://www.gbif.org/pt/occurrence/gallery?taxon_key=13
https://tgesbiogeek.weebly.com/pteridophyta.html
http://universe-review.ca/R10-34-anatomy203.htm
https://professores.unisanta.br/maramagenta/pteridofitas.asp
http://www.anatomiavegetal.ib.ufu.br/Xilema_texto.htm
https://tentativeplantscientist.wordpress.com/tag/pteridophyta/
https://tentativeplantscientist.wordpress.com/2013/05/07/plant-divisions-lycopodiophyta/
https://www.uwgb.edu/biodiversity/herbarium/pteridophytes/fern_intro01.htm
https://www.coursehero.com/sg/introduction-to-biology/life-cycle-of-pteridophytes/
https://courses.botany.wisc.edu/botany_401/lecture/03Lecture.html
https://ucmp.berkeley.edu/seedplants/cycadophyta/cycads.html
https://www3.nd.edu/~fboze/cycads.shtml
https://www.britannica.com/plant/sago-palm
http://www.biologyreference.com/Mo-Nu/Monocots.html
https://bio.libretexts.org/Bookshelves/Introductory_and_General_Biology/Book%3A_General_Biology_(Boundless)/30%3A_Plant_Form_and_Physiology/30.2%3A_Stems/30.2C%3A_Primary_and_Secondary_Growth_in_Stems
https://scied.ucar.edu/tree-rings
http://www.gradadm.ifsc.usp.br/dados/20161/SLC0622-1/aula%205%20parte1.pdf
https://www.meucantinhoverde.com/2012/05/tulipa-tulipa-hybrida.html
https://biologydictionary.net/dicot/
https://bio.libretexts.org/Bookshelves/Introductory_and_General_Biology/Book%3A_General_Biology_(Boundless)/32%3A_Plant_Reproductive_Development_and_Structure/32.2%3A_Pollination_and_Fertilization/32.2E%3A_Development_of_the_Seed
https://www.sparknotes.com/biology/plants/plantstructures/section1/
https://www.mundovestibular.com.br/estudos/biologia/fruto-e-pseudofrutos
http://www.anatomiavegetal.ib.ufu.br/pdf-recursos-didaticos/morfvegetalorgaFRUTO.pdf
http://segundocientista.blogspot.com/2015/09/botanica-scientia-amabilis.html
https://www.sobiologia.com.br/conteudos/bioquimica/bioquimica10.php
https://www.infoescola.com/citologia/tilacoide/
https://www.nature.com/scitable/topicpage/plant-cells-chloroplasts-and-cell-walls-14053956/
https://courses.lumenlearning.com/wm-biology1/chapter/reading-unique-features-of-plant-cells/
https://ssec.si.edu/stemvisions-blog/what-photosynthesis
https://www.biology-pages.info/G/GasExchange.html
https://www.sparknotes.com/biology/plants/plantstructures/section4/
http://www.webexhibits.org/causesofcolor/7A.html
https://www.sobiologia.com.br/conteudos/bioquimica/bioquimica14.php
https://www.sobiologia.com.br/conteudos/bioquimica/bioquimica15.php
http://educacao.globo.com/biologia/assunto/fisiologia-celular/fotossintese.html
https://www.sobiologia.com.br/conteudos/bioquimica/bioquimica16.php
https://www.peoi.org/Courses/Coursesen/bot/temp/bot16t91.html
https://sciencing.com/how-do-nonvascular-plants-get-water-nutrients-13428005.html
http://www3.botany.ubc.ca/bryophyte/mossintro.html
http://www.criptogamas.ib.ufu.br/node/546
https://courses.lumenlearning.com/wm-biology2/chapter/bryophytes/
http://universe-review.ca/R10-34-anatomy202.htm
https://bio.libretexts.org/Bookshelves/Introductory_and_General_Biology/Book%3A_General_Biology_(Boundless)/25%3A_Seedless_Plants/25.3%3A_Bryophytes/25.3B%3A_Liverworts_and_Hornworts
http://plantphys.info/plant_biology/lechtml/hornwort/
https://cronodon.com/BioTech/liverworts.html
https://www.britannica.com/topic/list-of-mosses-2075729
https://www.gbif.org/pt/occurrence/gallery?taxon_key=13
https://tgesbiogeek.weebly.com/pteridophyta.html
http://universe-review.ca/R10-34-anatomy203.htm
https://professores.unisanta.br/maramagenta/pteridofitas.asp
http://www.anatomiavegetal.ib.ufu.br/Xilema_texto.htm
https://tentativeplantscientist.wordpress.com/tag/pteridophyta/
https://tentativeplantscientist.wordpress.com/2013/05/07/plant-divisions-lycopodiophyta/
https://www.uwgb.edu/biodiversity/herbarium/pteridophytes/fern_intro01.htm
https://www.coursehero.com/sg/introduction-to-biology/life-cycle-of-pteridophytes/
https://courses.botany.wisc.edu/botany_401/lecture/03Lecture.html
https://ucmp.berkeley.edu/seedplants/cycadophyta/cycads.html
https://www3.nd.edu/~fboze/cycads.shtml
https://www.britannica.com/plant/sago-palm
http://www.biologyreference.com/Mo-Nu/Monocots.html
https://bio.libretexts.org/Bookshelves/Introductory_and_General_Biology/Book%3A_General_Biology_(Boundless)/30%3A_Plant_Form_and_Physiology/30.2%3A_Stems/30.2C%3A_Primary_and_Secondary_Growth_in_Stems
https://scied.ucar.edu/tree-rings
http://www.gradadm.ifsc.usp.br/dados/20161/SLC0622-1/aula%205%20parte1.pdf
https://www.meucantinhoverde.com/2012/05/tulipa-tulipa-hybrida.html
https://biologydictionary.net/dicot/
https://bio.libretexts.org/Bookshelves/Introductory_and_General_Biology/Book%3A_General_Biology_(Boundless)/32%3A_Plant_Reproductive_Development_and_Structure/32.2%3A_Pollination_and_Fertilization/32.2E%3A_Development_of_the_Seed
https://www.sparknotes.com/biology/plants/plantstructures/section1/
https://www.mundovestibular.com.br/estudos/biologia/fruto-e-pseudofrutos
http://www.anatomiavegetal.ib.ufu.br/pdf-recursos-didaticos/morfvegetalorgaFRUTO.pdf
https://sistemasdeproducao.cnptia.embrapa.br/FontesHTML/Tomate/TomateIndustrial_2ed/deficiencias.htm
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.